About my work (sex, heart, muscles, and the fruit-fly)
 The field I work in is Developmental Biology -- a discipline that attempts to investigate how all the strikingly different and specialized cells making up a complex living organism come into being. This is a subtler question than might appear at first glance: for, in almost all cases, all the cells in an organism contain the same genetic blueprint, DNA, yet end up choosing different fates. Understanding the manner in which this is achieved -- the signals and mechanisms used -- is the primary goal in developmental biology.
The field I work in is Developmental Biology -- a discipline that attempts to investigate how all the strikingly different and specialized cells making up a complex living organism come into being. This is a subtler question than might appear at first glance: for, in almost all cases, all the cells in an organism contain the same genetic blueprint, DNA, yet end up choosing different fates. Understanding the manner in which this is achieved -- the signals and mechanisms used -- is the primary goal in developmental biology.
My previous work (as a graduate student completing his PhD) dealt with certain aspects of sex determination and sexual differentiation in the fruit-fly Drosophila melanogaster. My current research is focused on the genes involved in the proper development of the heart and muscles using Drosophila once again as a model system. A brief description of both these research projects is presented below.
Sex and the fruit-fly
With a title like that, I suspect the reader cannot help but ask the following questions:
- Why study sex?
- Why study it in Drosophila?
- And what particular aspects of sex in Drosophila did I focus on, anyway?
Let me try to address these questions one at a time.
Why study sex?
Leaving aside the glib retort that sex is a phenomenon that is of intrinsic interest to all human beings, one has to understand that, from an evolutionary perspective, sex is an absolutely wonderful innovation. The far more efficient system of amassing advantageous mutations that it brings into play allows a sexual species to adapt to changes in its environment much more easily than an asexual species. It should thus come as no surprise to realize that sex has evolved independently more than once, very many times in fact.
And in all these cases of independent evolution, the mechanisms by which sex is determined have, naturally, been quite different. In mammals like us, for instance, it is the presence or absence of the Y-chromosome that determines whether a particular organism is male or female. In ants and bees, males are the individuals that are produced parthenogenetically, that is, without fertilization; females are those that emerge from fertilized eggs. In some species of reptiles like turtles, lizards, and crocodiles, sex is determined by the temperature at which the eggs develop. And there are schools of fish in which the largest animal is the male; if he were to meet an untimely end, the largest female in the school would gradually become transformed into a male. Similarly, in the marine worm Bonellia, young worms are indeterminate sexually, but are attracted to females. If they can attach themselves to a female, they become transformed into males and live inside the female body as a parasite. If they are unable to attach themselves to any female, they develop into females themselves. Obviously all these systems present the developmental biologist with an absolute plethora of mechanisms by which the fates of cells become determined -- making the study of sex a very promising undertaking indeed.
Why study sex in Drosophila?
A simple response might well be: for the very same reasons that we pursue so many other biological issues in Drosophila. This answer is not as flippant as it might first appear; for reasons that are described in more detail in Gerard Manning's Introduction to Drosophila in the WWW Virtual Library, this organism is an almost perfect model system for addressing biological questions. First, it has been used as an experimental system for so very long that considerably more is known about it (compared to other systems) and very many tools and techniques have been developed -- making it both easy to handle and very well understood. Its small size and its short life cycle make it cheap and easy to keep in large numbers. Along with the ease of mutagenesis, this makes it particularly amenable to genetic analyses -- not an overly easy task in a number of other systems. Finally, many Drosophila genes have been found to have counterparts in Homo sapiens, suggesting that some of our understanding of Drosophila biology might also be applicable to human beings.
There remains, however, a more specific reason for studying sex determination in Drosophila: the fact that it uses quite a novel and interesting scheme. Sex in Drosophila is not brought about by the presence or absence of the Y-chromosome (though generally males have one while females lack any); no, it is instead the ratio of the X-chromosomes to the other chromosomes (autosomes) that determines sex. When the ratio is 1:1, that is, the individual contains two X-chromosomes and a pair of each autosome, the fly is female; when the ratio is 1:2 (one X-chromosome and a pair of each autosome), the fly is male.
This entire process of sex determination is mediated through a gene called Sex-lethal that acts as a master switch. In response to the appropriate X-chromosome:autosome ratio, the gene remains in either an active or an inactive state, leading respectively to either female or male development. In both cases, Sex-lethal lies at the head of a hierarchy of genes, a hierarchy that gradually branches out to control different aspects of sex: dosage compensation (the mechanism that ensures the same amount of RNA and thus protein is made despite the female fruit-fly having twice as many X-chromosomes), sexual behaviour, germline sex differentiation (sex determination of the cells that will ultimately give rise to sperm or eggs), and somatic sex differentiation (sex determination of all other cells).
The study of sex in Drosophila revolves around indentifying all the agents that are involved in this hierarchy (more and more are gradually being discovered) and investigating the mechanisms by which they interact.
What particular aspects of sex in Drosophila did I focus on?
The reader who has glanced at the sex determination hierarchy might have noticed that male and female sexual differentiation in the somatic cells is brought about by the action of the doublesex gene. The products of the doublesex gene are transcription factors, proteins that regulate where and when other genes are turned on.
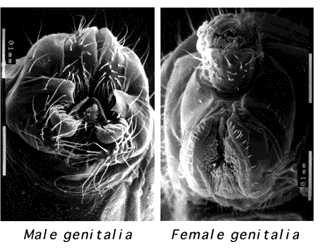 It is these other genes, direct or indirect targets of doublesex, that I was interested in. doublesex and all the genes above it in the hierarchy function essentially as regulatory switches, serving to fine-tune and transmit the X-chromosome:autosome sex determining signal along different pathways. Unlike them, I expected these target genes to be the actual effectors of sex determination, the genes which directly bring about one or more particular sex-specific fates. I decided to look for these genes in the genital imaginal disc, the precursor to the structures that show the most extreme degree of sexual dimorphism in flies -- the male and female genitalia and anal structures.
It is these other genes, direct or indirect targets of doublesex, that I was interested in. doublesex and all the genes above it in the hierarchy function essentially as regulatory switches, serving to fine-tune and transmit the X-chromosome:autosome sex determining signal along different pathways. Unlike them, I expected these target genes to be the actual effectors of sex determination, the genes which directly bring about one or more particular sex-specific fates. I decided to look for these genes in the genital imaginal disc, the precursor to the structures that show the most extreme degree of sexual dimorphism in flies -- the male and female genitalia and anal structures.
My project involved identifying these target genes and understanding both how they are utilised to bring about sexual differentiation and how they are regulated by both the sex determination hierarchy and other hierarchies (for while the sex determination hierarchy will determine whether they are on in say, males or in females, clearly it is the influence of other hierarchies that will determine the spatial and temporal patterns in which they are expressed). Some of this work is described in more detail in the following papers (PDF format):
- Ahmad, S. M. and Baker, B. S.
- Sex-specific deployment of FGF signaling in Drosophila recruits mesodermal cells into the male genital imaginal disc. Cell 2002, 109: 651-661.
- Christiansen, A. E., Keisman, E. L., Ahmad, S. M. and Baker, B. S.
- Sex comes in from the cold. Trends in Genetics 2002, 18:510-516.
-
The heart and muscles
Forthcoming...
Return to:
Papers are in Adobe Acrobat (PDF) format. The Acrobat Reader available from the Free Adobe Acrobat Reader Software page will allow these papers to be viewed.
Scanning electron micrographs of male and female Drosophila genitalia were provided by Dr. Hao Li.
Revised: July 9, 2007